Subalpine and Alpine Wildflowers and Pollinators of the North Cascades: Part 2

By Lauren Ridder, graduate student in the institute’s 14th cohort. This is part two of Lauren’s natural history project. Find part one here.
The Biome: Subalpine to Alpine
The alpine lifezone or biome is most often described as the area above or near treeline on mountaintops. In the North Cascades, the elevation range of the alpine zone is from about 6,400ft to about 8,530ft (Douglas & Bliss, 1977). The subalpine biome often shares many characteristics with alpine plant and animal communities as the boundaries between the two lifezones are rather indistinct (Billings, 1974). The varying topography blends these two biomes, making the assignment of plant communities highly subjective. Among the main features that designate an area as subalpine are the discontinuation of the forest and the formation of “scattered tree clumps in a meadow mosaic” (Taylor & Douglas, 1995, p.4). These tree clumps are pioneers in harsh soil and growth conditions and “are normally short, with spreading branches, but [they still] retain definite crowns and do not develop the dense, low, thicketlike growth form known as krummholz” (Taylor & Douglas, 1995, p. 5-6). Krummholz is generally found on higher slopes and marks the beginning of the alpine zone (Taylor & Douglas, 1995). It would be much easier to assign general characteristics to these biomes if the mountain were flat ground with consistent weather patterns. However, nature provides large doses of interest and variety in vegetation patterns through vastly different slope aspects, substrate conditions, and extreme daily ranges in temperature, wind speeds, solar radiation, and water availability.
The combination of these abiotic factors creates many different habitats with microclimates and vegetation stripe communities occurring within those habitats. Fellfields are most common in the alpine biome, and are “characterized by rocky ground and dry soil, and are typically less than half covered by vegetation…Plants that grow here must be short in stature or they will be desiccated by freezing wind in winter and blasted by wind-driven sand in summer” (Visalli, 2014b, p.3). The constant frost action and avalanche potential acting on the slopes causes the soil to be unstable, poorly developed, and easily eroded as well, and pioneering plants must act quickly to establish roots when possible (Douglas & Bliss, 1977). Vegetation stripes can occur in the talus and scree slopes of fell- and boulder-fields, where soil and moisture has found a path of least resistance to percolate through or flow down and created a pocket of nutrients for plants to capitalize on (Douglas & Bliss, 1977).
Wildflowers are more commonly found in the moist meadows and wetlands of the subalpine and low alpine biomes. Here the plants have better access to water since drainage is poor, and they are more protected from wind, so they can grow taller to “[compete] for sunlight and for the attention of pollinators” (Visalli, 2014b, p.3). The location of these meadows on the mountain can have an effect on the types of plants that are successful as well. For example, southern facing slopes have longer access to water as snow melts and glacial runoff contributes to the water table, but an exposed meadow is basically in the midst of a drought come July and August. On the other side of the mountain, snow melts later on the northern slopes, and plants must instead deal with a short growing season (Taylor & Douglas, 1995). The timing of snowmelt is the main environmental factor that determines the makeup of the plant communities found on mountain slopes.
Physical Adaptations to the Biome
Over time, the challenging mountaintop environment has led to the development of certain morphological characteristics that enable a plant to be successful in the alpine and subalpine biomes. In the alpine, plants tend to have a short, cushion-like growth form, while in the lower, wetter subalpine meadows, plants can grow taller and more structurally complex. The strong winds along ridgelines and exposed slopes flow “over the streamlined cushion much as over an airplane wing, and the form provides the maximum photosynthetic surface with the minimum exposure to physical damage” (Pojar & MacKinnon, 2013, p.135). The low form also concentrates warmth and solar radiation at ground level, which warms the roots and creates a more favorable microclimate within a microclimate (Taylor & Douglas, 1995). Most alpine and subalpine vegetation is also covered in fine, silky hairs that not only diffuses the harsh alpine light, but also “insulates them from rapid temperature change and reduces water loss on hot, windy summer days” (Taylor & Douglas, 1995, p.7). Deep pigmentation and rich colors on the leaves and flowers assists with ramping up metabolism after chilly nights, as well as acting as a sunscreen that protects vulnerable new growth tissue (Pojar & MacKinnon, 2013). Beneath the surface of the soil, alpine and subalpine plants have found success in the development of extensive root systems. These root systems allow for soil build-up, erosion control, and carbohydrate storage for use in the following growth season. In fact, “much of the first year’s photosynthate goes into the development of a root system” in order to successfully establish the plant (Billings, 1974, p.134). The aboveground and below-the-surface features allow plants to take full advantage of the minimal resources available to them in an unforgiving and ever-changing environment. These physical adaptations took thousands of years to evolve however, and the rate of climate change will quickly surpass the rate at which specialized alpine and subalpine plants can adapt. “Most plant species cannot move fast enough to keep up with the projected changes,” which will result in a loss of diversity and a shift in what alpine and subalpine meadows look like in the coming decades (Pojar & MacKinnon, 2013, p.36).
Co-evolution with Pollinators
The symbiotic relationship that has evolved between angiosperms and their pollinators is fascinating and incredibly complex. The extent to which each individual species has tailored their morphology and behavior to fulfill basic reproductive and nutrient needs is astounding. While overall patterns of wildflower and pollinator relationships will be focused on, it is important to remember that this is a generalized description of the connections between plant and pollinator. Oversimplification is a slippery slope and while it can help researchers to understand the processes driving evolution, there are so many other contributing factors that have yet to be discovered, let alone described.
Many of the plants found at higher elevations are generalists, meaning they take advantage of many different pollinators’ behaviors rather than focus morphologically on one species. Too specialized of a relationship between plant and pollinator would be detrimental, especially in this age of climate change. Co-evolution began as a reward system, of course, and plants can be very rewarding. The main benefits that pollinators receive from wildflowers are pollen and nectar, though plants have also adapted to provide “nutritious tissues, oils, waxes, resins and perfumes, with the understanding that a given pollinator is not interested in all of these, but concentrates on just one or two” (Meeuse & Morris, 1984, p.31). Pollen is one of nature’s super foods and contains “protein, sizeable amounts of starch, sugars, fat or oil, minerals, antioxidants, and vitamins such as thiamin. It is also rich in free amino acids” (Meeuse & Morris, 1984, p.31). This combination makes pollen an irresistible and easily accessed nutrient source for a variety of insect pollinators.
Nectar is also perfectly packaged and poised for use by pollinators, as it is “easily manufactured; it can be produced in controlled quantities as demand requires; animal visitors can gather it simply by sucking up the fluid, which takes little time and effort; and as a solution of readily digested sugars, it is a quickly assimilated source of energy” (Meeuse & Morris, 1984, p. 34). Nectar is such an important fuel, that it may have been produced before flowers had even evolved. Nectaries are present on more ancient plant groups such as ferns as a fuel source for young leaves and other building materials. Excess nectar may have been excreted, so it was only a matter of time before an ancient pollinator found its way to the nectary, a wayward spore was carried to a neighboring fern, and a beautiful and complex relationship was born (Meeuse & Morris, 1984). Over time, plants found that nectar could be modified to attract certain types of pollinators as well; for example, birds cannot ingest concentrated, syrupy nectar, so bird-pollinated flowers produce a less concentrated solution. The amount can be adjusted, too. Ant-pollinated plants will only make a small amount of nectar to discourage other, larger and potentially more destructive pollinator visits (Meeuse & Morris, 1984).
Plants have developed several methods to advertise these superfoods and fuels to would-be pollinators. As extremely visual creatures, color is usually the wildflower characteristic that humans notice first. The same holds true for insects, who have excellent eyesight and many of which can also see in the ultraviolet (UV) spectrum. The UV spectrum plays an important part in guiding pollinators towards the nectar or pollen payload. Many flowers have distinct nectar guides that are invisible to humans unless viewed through a UV filter. For example, what may appear to humans as a slight variation of yellow in the center of a wild buttercup is actually a clear target directing pollinators to the flower’s anthers.

A primrose illuminated by ultraviolet light reveals the nectar guides used by pollinators that are normally hidden from a human’s eye. Source: Bjorn Roslett, Science Photo Library
A flower’s color is produced through varying amounts of pigments arranged side by side, in different cells, combined in a single cell, or superimposed on each other. The hue can be modified by the shape of the cell that the pigments are in, the presence of air spaces, or by waxy film or hair on the surface of the plant structure. The pigments can also change color over time- after pollination or with age for example- simply by changing the acidity of the fluid in the cell (Meeuse & Morris, 1984). One pigment often contained in new alpine plant growth is a “red-colored carbohydrate pigment called anthocyanin, which acts both as a block to harmful ultraviolet light (which is more intense at high altitudes than in the lowlands and can damage plant cells just as it can damage human cells), and is a more effective heat absorber than the chlorophyll-green coloration that follows in later growth stages” (Visalli, 2014b, p.11). Another type of pigment usually present in subalpine and alpine wildflowers is the anthoxanthins, which create a pale ivory to deep yellow color. Wildflowers with these pigments may appear to be ivory white to humans, but they will “absorb ultraviolet light very strongly and are therefore perceived as colored (blue-green) by insects such as honey-bees” (Meeuse & Morris, 1984, p.25). In some daisies, a white color is achieved through air pockets between cells. The air space and resulting refraction of light is also the reason we perceive snow to be white (Meeuse & Morris, 1984).
Color is not the only wildflower attribute that brings in pollinators, nor is it the most effective. Scent is a powerful motivator towards building flower constancy, or “the ‘loyalty’ which a pollinator displays towards the flowers of just one plant species. The pollinator is perfectly capable of pollinating other flower species, but will stick to its original behavioral pattern even when flowers that have more to offer in terms of nectar and pollen are becoming available on the landscape” (Meeuse & Morris, 1984, p.31). This is a learned behavior pattern that promotes efficiency in pollination for both the pollinator and the plant. Many insects have well developed senses of smell, so fragrance is key is attracting the right pollinator for the job at the right time. Wildflowers are capable of adjusting the potency of their scent throughout the day in order to signal to pollinators when flowers will be open. This enables honeybees and bumblebees, who have strong flower constancy tendencies, to adhere to a strict pollination schedule for a few plants a season (Meeuse & Morris, 1984). Signaling prime pollination time through scent also helps some wildflowers avoid self-pollination. For example, the peak time for collection of Fireweed (Epilobium angustifolium) pollen is around one o’clock in the afternoon when the blooms near the bottom of the flower are open and the anthers are releasing pollen. To prevent self-pollination the anthers have matured before the stigmas on the same plant are receptive to pollen- a practice called protandry (Meeuse & Morris, 1984). All of these things working together effectively advertise to pollinators the brief window of time available for pollination in the alpine and subalpine biomes season after season.
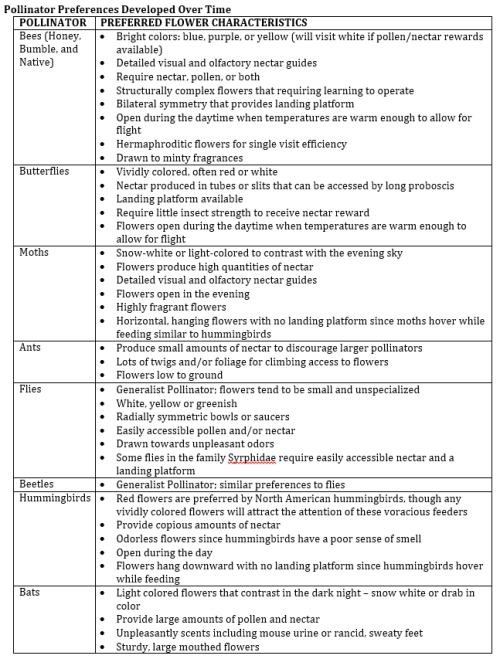
Pollinator Preferences Developed Over Time
Wildflowers and The Effects of Climate Change
The morphology of wildflowers took thousands of years to evolve, and the rate of climate change will quickly surpass the rate at which the highly specialized alpine and subalpine plants can adapt. These plants are already well adapted to a harsh environment and have no way of relocation when the situation changes. A global increase in temperature impacts snowmelt timing- the most influential of environmental factors on subalpine wildflowers. The brilliantly colored Indian paintbrush (Castilleja miniata) is already seeing the effects of altered seasonal temperatures. The coloration and sturdiness of the flower’s bracts attract butterflies, specifically the Anicia checkerspot butterfly. A recent study found that the paintbrush’s growing season is 9.5 days shorter in dry soils compared to moist soils, “so as the climate warms and soil becomes drier, the red paintbrush will experience shorter growing periods, which will in turn affect the Anicia checkerspot, who thrive on the plant” (National Park Service, n.d.). Another iconic North Cascadian wildflower, the yellow glacier-lily (Erythronium grandiflorum), has been impacted by climate change. Their coloration and shape means pollination by moths and bees, though this delicate flower also has a relationship with a much larger creature, the Grizzly Bear. Glacier-lily bulbs are a great source of starch and fiber and are a stable of the grizzly’s diet. The bears ‘cultivate’ the bulbs when they dig up the bulbs and introduce nitrogen and ammonium to the soil, creating areas rich in nutrients for lily seedlings to establish. However, as the date of snowmelt advances year by year, so does the lily’s growing season, which has already begun to influence its relationship with its bumblebee pollinators (Pojar & MacKinnon, 2013). A decline in pollination opportunities means a decline in sexual reproduction and genetic diversity as well. If temperatures and precipitation amounts continue to fluctuate season after season, the mountaintops of Cascadia will look very different in the near future. The connections between plant, animal, insect, soil, water, and air will falter, and the beauty of a subalpine meadow in full bloom will become a distant memory.
References
Billings, W.D. (1974). Adaptations and Origins of Alpine Plants. Artic and Alpine Research, 6(2), 129-142.
Douglas, G.W., & Bliss, L.C. (1977). Alpine and High Subalpine Plant Communities of the North Cascades Range, Washington and British Columbia. Ecological Monographs, 47, 113-150.
Guertin, P., Barnett, L., Denny, E.G., Schaffer, S.N. (2015). USA National Phenology Network Botany Primer. USA-NPN Education and Engagement Series 2015-001. www.usanpn.org.
Meeuse, B., & Morris, S. (1984). The Sex Life of Flowers. New York, NY: Facts on File Publications.
National Park Service. (no date). Changing Plant Life. Natural Notes. Retrieved from http://www.nps.gov/noca/learn/nature/upload/NaturalNotes_07page3.pdf
Pojar, J., & MacKinnon, A. (1994). Plants of the Pacific Northwest Coast: Washington, Oregon, British Columbia, & Alaska. Vancouver, British Columbia, Canada: Lone Pine Publishing.
Pojar, J., & MacKinnon, A. (2013). Alpine Plants of the Northwest: Wyoming to Alaska. Edmonton, Alberta, Canada: Lone Pine Publishing.
Taylor, R.J., & Douglas, G.W. (1995). Mountain Plants of the Pacific Northwest. Missoula, MT: Mountain Press Publishing Company.
Visalli, D. (no date). A Key to the Common Flowing Plant Families of the Methow. The Methow Naturalist. http://methownaturalist.com/16- Key%20to%20Flowering%20Plants.PDF
Visalli, D. (2014a, July). Flora of the Methow: The Plants of the Methow in Their Evolutionary Context. Methow Naturalist. http://methownaturalist.com/7-2014-Flora%20of%20the%20Methow.pdf
Visalli, D. (2014b, Summer). Life in the High Country: A Naturalist’s Guide. The Methow Naturalist, 19(2), 1-3, 10-13. http://methownaturalist.com_2-MNv19n2web.pdf